







 Eusthenopteron (fish) → Tiktaalik (fish) → Acanthostega (tetrapod)
Eusthenopteron (fish) → Tiktaalik (fish) → Acanthostega (tetrapod)The transformation of fish into the kind of amphibians we see in the mid Palaeozoic is not inconceivable and probably happened. However, it is difficult to conceive of its happening by any random process. Water and air are very different environments. While water is liquid H2O, the oxygen component is unavailable for life; fish depend on the oxygen separately dissolved in the water, which varies with temperature and depth. At a temperature of 15 ?C close to the surface, it is around 10 milligrams per liter, or 1 part per 100,000. The concentration in air is 23 parts per 100. In addition, animals are buoyant in water because they mainly consist of water themselves; on land they are subject to gravity and motion requires more energy. They need legs to move around, or muscular scales if they have lost their legs. A shift from water to air therefore involves a radical re-organisation of embryonic development and adult design.
Modern theory prefers to designate the new animals as ‘tetrapods’ rather than amphibians – vertebrates with limbs and feet. That is because, contrary to the expected story, it has become apparent that the first four-footed animals acquired their feet while living entirely in the water. So tetrapods include newts and salamanders as well as exclusively terrestrial animals. The term gives the impression that all animals with four limbs have a common ancestor, in the same way as ‘arthropod’ implies that all invertebrates with segmented exoskeletons and legs have a common ancestor. But as with all beliefs about the past, that has to be demonstrated on the facts.
By the time the first tetrapod fossils appear, nine-tenths of Earth’s preserved geological time has already passed, and of that nine-tenths fish fossils occur only in the last twenty-fifth. As with all animal classes and phyla, the origins of fish are obscure. Even if we confine ourselves to relationships within the group, fishes take so many different forms that untangling their relationships is a challenge, and taxonomic schemes have changed over the years. Currently, living fish are divided into 5 classes:

- Hagfish (Myxini)
- Lampreys (Petromyzontida)
- Cartilaginous fish (Chondrichthyes)
- Lobe-finned fish (Sarcopterygii)
- Ray-finned fish (Actinopterygii)
The last two are ‘bony fish’ (Osteichthyes), distinct from cartilaginous, and may have had a common ancestor. The exact number is not crucial, so we will assume that they did, and that otherwise these classes each stem from a single ancestor. Thus four lineages account for all the fish in the seas today. That is of course a vast amount of evolution, and of a most wonderful kind. Whether the four classes were themselves interrelated is impossible to test because the fossil material prior to their appearance is too poor.
Classes subdivide into orders. Hagfish and lamprey each consist of only 1 order. Cartilaginous fish comprise 14: chimaeras, sharks of various kinds, skates, sawfish and rays. Lobe-finned fish consist of 2 orders: coelacanths and lungfish. Ray-finned fish comprise over 40 and now encompass the vast majority of fish in the sea, from seahorses to eels, being more diverse in number of species (there are over 30,000) than all mammals, birds and reptiles put together. Astonishingly, most of that diversity arose late in geological time, during the Late Cretaceous and Cenozoic, when oceans began to cool and deepen. The delay, followed by such a dazzling display of creativity, suggests that they were programmed to diversify in response to the cooling.
- Conodonts
- Pteraspidomorphs (including the heterostracans)
- Anaspida
- Osteostracans
- Placoderms
- Acanthodians
Acanthodians probably descended from fish ancestral to the cartilaginous group (Coates et al. 2018). The general evolutionary scheme scheme, supported by the fossil record, is that fish were initially finless and jawless, gradually evolving fins and jaws as competition for resources grew. In that case, fins and jaws evolved independently in the different lineages.
From earliest to latest, the first occurrences of these eleven classes, some living, some extinct, are as follows (dates in millions of years, radioisotope chronology):
| Class | Period | Date | Type | Genus |
| Conodonts | Cambrian | 500 | jawless | various |
| Pteraspidomorphs | Ordovician | 480 | jawless | Arandaspis |
| Osteostracans | Silurian | 440 | jawless | Ateleaspis |
| Acanthodians | Silurian | 440 | jawed | various |
| Anaspids | Silurian | 435 | jawless | Pharyngolepis |
| Lobe-finned osteichthyans | Silurian | 420 | jawed | Guiyu |
| Placoderms | Devonian | 416 | jawed | various |
| Cartilaginous fish | Devonian | 409 | jawed | Doliodus |
| Ray-finned osteichthyans | Devonian | 390 | jawed | Cheirolepis |
| Lampreys | Devonian | 365 | jawless | Priscomyzon |
| Hagfish | Carboniferous | 300 | jawless | Myxinikela |
The existence of more classes in the past than in the present also does not conform to evolution theory. As has been discussed in relation to the Cambrian Explosion, Darwinism predicts that morphological disparity should increase with time.
The general pattern is that fish fossils were initially rare, reflecting a world where fish were few and far between. The colonising fauna came from the open sea, where the floors beneath them were hot, basaltic wastelands. Since the only plentiful food available was algae and microscopic plankton, they had no use for teeth and jaws. Only as near-shore environments became more habitable did they acquire equipment enabling them to nibble seaweeds, filter sediment, crack open shells and capture other fish. It wasn’t a process of natural magic; they were programmed to do so, according to their kinds. As the available niches became more various, as environments began to be vegetated and to host a range of invertebrate life, so the fish moving into them became more specialised.
Eusthenopteron is sometimes figured as crawling out of the mud of a Devonian lake, apparently with the intention of finding another pool to swim in. Because Eusthenopteron was once cast in the role of the ‘ancestor’ of tetrapods, tetrapodlike behavior was attributed to it. However, taking the whole morphology of the fish, with its streamlined torpedo shape, and dorsal, anal, and pelvic fins placed near the back of the body, it seems that the lifestyle of Eusthenopteron was much more like that of a modern pike (Esox), a fully aquatic lurking predator.
Jennifer Clack, Gaining Ground, p. 63 (2002)
In the absence of better candidates, Eusthenopteron was was regarded as well on the way to becoming a tetrapod, and was portrayed as such in illustrations. However, it has since been relegated to a side branch. So has its contemporary, Panderichthys. ‘Palaeontologists didn’t previously have a decent fossil representing the intermediate between finned fish and four-footed land animals,’ admitted Bob Holmes in the New Scientist. But the admission was made only after the discovery of Tiktaalik.
 This fossil, a large Late-Devonian fish, undoubtedly narrowed the gap between the fish Panderichthys and the tetrapod Acanthostega. As soon as the embargo on the scientific report was released, press and radio promulgated the news across the world. The Guardian went so far as to say it was one of the most important fossil finds in history: a missing link between fish and land animals showing how creatures first walked out of water and onto dry land. Nature itself was scarcely more phlegmatic. In the context-setting commentary that accompanied the report Jennifer Clack and Per Ahlberg said they expected it to acquire the same iconic status as Archaeopteryx. Was the hype justified?
This fossil, a large Late-Devonian fish, undoubtedly narrowed the gap between the fish Panderichthys and the tetrapod Acanthostega. As soon as the embargo on the scientific report was released, press and radio promulgated the news across the world. The Guardian went so far as to say it was one of the most important fossil finds in history: a missing link between fish and land animals showing how creatures first walked out of water and onto dry land. Nature itself was scarcely more phlegmatic. In the context-setting commentary that accompanied the report Jennifer Clack and Per Ahlberg said they expected it to acquire the same iconic status as Archaeopteryx. Was the hype justified?The relationship of limbed vertebrates (tetrapods) to lobe-finned fish (sarcopterygians) is well established, but the origin of major tetrapod features has remained obscure for lack of fossils that document the sequence of evolutionary changes. Here we report the discovery of a well-preserved species of fossil sarcopterygian fish from the Late Devonian of Arctic Canada that represents an intermediate between fish with fins and tetrapods with limbs, and provides unique insights into how and in what order important tetrapod characters arose. Although the body scales, fin rays, lower jaw and palate are comparable to those in more primitive sarcopterygians, the new species also has a shortened skull roof, a modified ear region, a mobile neck, a functional wrist joint, and other features that presage tetrapod conditions.
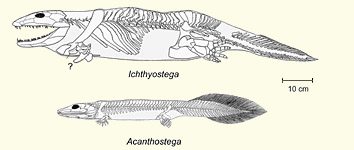
 Tiktaalik was a lobe-finned fish, in the same broad group as coelacanths and lungfish, living at a time when lobe-fins were far more diverse than today. The new genus/species adds to that diversity. Although still a fish, Tiktaalik does allow itself to be interpreted as an animal transitional to the earliest fossilised aquatic or semi-aquatic tetrapods. Shared features include: a lengthened snout (measured from the eyes to the tip of the skull), a mobile neck (facilitated by the loss of certain bones at the back of the skull), overlapping (‘imbricate’) ribs and a pectoral girdle that may have given it an ability to lift the front part of its body by its fins. Stratigraphically, it appears after Pandericthys and before the first four-limbed animals, Acanthostega and Ichthyostega, though the gap is only (in Darwinian terms) around 5 million years. Since these two tetrapods were very different from each other, presumably their Tiktaalik-like ancestor evolved in two separate directions, with Acanthostega on one branch and Ichthyostega on the other.
Tiktaalik was a lobe-finned fish, in the same broad group as coelacanths and lungfish, living at a time when lobe-fins were far more diverse than today. The new genus/species adds to that diversity. Although still a fish, Tiktaalik does allow itself to be interpreted as an animal transitional to the earliest fossilised aquatic or semi-aquatic tetrapods. Shared features include: a lengthened snout (measured from the eyes to the tip of the skull), a mobile neck (facilitated by the loss of certain bones at the back of the skull), overlapping (‘imbricate’) ribs and a pectoral girdle that may have given it an ability to lift the front part of its body by its fins. Stratigraphically, it appears after Pandericthys and before the first four-limbed animals, Acanthostega and Ichthyostega, though the gap is only (in Darwinian terms) around 5 million years. Since these two tetrapods were very different from each other, presumably their Tiktaalik-like ancestor evolved in two separate directions, with Acanthostega on one branch and Ichthyostega on the other.
Imbricate ribs do not occur in any other fish, nor in Acanthostega, but do occur in Ichthyostega. Since even that animal was primarily aquatic, they are unlikely to have played a role in supporting the weight of the animal. The stiffening of the spine produced by this arrangement would have inhibited horizontal flexion (the wiggling motion of most fishes) just as it would have done in Ichthyostega (Clack 2006), and this fact alone is sufficient to explain the mobile neck and fin joints: without some compensating mobility the animal would not have been viable as a predator. In overall morphology, the ribcages of Tiktaalik and Ichthyostega are unlike each other.
{kind=link}
{kind=link}
A Devonian rhizodontid, Sauripterus, is known to possess digit-like radials, but phylogenetic analyses indicate that this group is not the closest relative of tetrapods.
The pelvic fin was not capable of bearing significant weight. In contrast to both tetrapods and lungfish, pelvic girdle’s two halves were unfused and they were not attached to the spine, except possibly by unfossilised ligaments. On the other hand, whereas in Panderichthys the pelvic girdle was smaller than the pectoral girdle, in Tiktaalik their size was about the same. The corresponding fins may or may not have been similar in size: the preserved lepidotrichia suggest that they were, whereas the pectoral fins of Elpistostege, Tiktaalik’s closest relative, were three times bigger than the pelvic fins. In the early tetrapods the hindlimbs were bigger and more powerful than the forelimbs.
. Gill cover in blue, postparietal bones in green.") The animal does not fall plumb in the middle between Panderichthys and Acanthostega. It is a fish, albeit an unusual one, and while there is only a relatively small gap between Panderichthys and Tiktaalik, a big gap between Tiktaalik and Acanthostega remains. In a significant number of features, body scales, fin rays, lower jaw, palate, the fossil resembles sarcopterygians that are considered to be evolutionarily less advanced. As Ahlberg and Clack say, ‘we have almost no information about the step between Tiktaalik and the earliest tetrapods, when the anatomy underwent the most drastic changes.’ It may be reasonable enough to link the first pair into a group that had a common ancestor (the ‘elpistostegalian fish’), but it requires a lot more extrapolation to link the second pair together.
The animal does not fall plumb in the middle between Panderichthys and Acanthostega. It is a fish, albeit an unusual one, and while there is only a relatively small gap between Panderichthys and Tiktaalik, a big gap between Tiktaalik and Acanthostega remains. In a significant number of features, body scales, fin rays, lower jaw, palate, the fossil resembles sarcopterygians that are considered to be evolutionarily less advanced. As Ahlberg and Clack say, ‘we have almost no information about the step between Tiktaalik and the earliest tetrapods, when the anatomy underwent the most drastic changes.’ It may be reasonable enough to link the first pair into a group that had a common ancestor (the ‘elpistostegalian fish’), but it requires a lot more extrapolation to link the second pair together.
There is, moreover, a large gap after Ichthyostega. It often fails to be mentioned that, so far as the fossil record is concerned, both Acanthostega and Ichthyostega are dead-ends. They shed no light on the origin of modern tetrapods, aquatic or terrestrial.
According to Nature‘s announcement of the fossil, Tiktaalik was a fish that crawled out of the water, but if so, the lineage must have crawled back again, for Acanthostega ‘rarely, if ever, made forays onto dry land and its legs would have almost certainly been incapable of supporting its body had it done so’ (Clack 2002, p 128f). That’s less than was claimed for this animal. We were told in one breath that Tiktaalik had a long snout that would have been suited to catching prey on land, in the next that it hauled itself onto land only to escape predators.
One of the monkfish was observed to move along the seabed for several metres at a time by walking. Several ‘walks’ were observed. The gait involved both the pelvic and pectoral fins and the body and tail were lifted clear of the seabed. The pelvic fins appeared to be the main weight bearing fins lifting the body up from the seabed. They also seemed to be responsible for a considerable proportion of the forwards propulsion.
Frogfish, handfish and monkfish are members of the order Lophiiformes or anglerfish, within the subclass of teleost fish, within the class of ray-finned fish. The evolutionary history of these walkers is obscure but we may presume that their particular forms are the result of immense diversification over time within the order. Most species inhabit relatively deep water. Certainly their appendages are quite unrelated to those of lobe-finned fish. Anglerfish are an example of how wonderfully unconventional life can be, as if whoever designed them was a non-conformist who delighted in subverting stereotypes.
 Thinking that was less ideologically driven might not go amiss in the present case. While locomotion may have been a function of Tiktaalik’s front fins, habitat indicated by the geological setting was a floodplain, dissected from time to time by channels. Other fossils recovered from the locality include placoderms and a variety of other lobe-fins. Out of water, Tiktaalik could, at best, only have dragged its 2½-metre-long body. Monkfish, which like Tiktaalik have a wide mouth, flattened skull and eyes on top of the skull rather than on the sides, again alert us to other possibilities, for in addition to walking they use their fins to scoop out sediment from under their bodies and make a hollow, in which, having camouflaged themselves to match their surroundings, they lie in wait. At the right moment, they strike the prey by suddenly pushing with their fins upwards and forwards. The feeding strategy of Tiktaalik may have been similar. Several lineages of fish, such as mudskippers, rockskippers and blennies, have even evolved an ability to crawl out of water. However, there is no reason to think they are on the way to leaving the water altogether.
Thinking that was less ideologically driven might not go amiss in the present case. While locomotion may have been a function of Tiktaalik’s front fins, habitat indicated by the geological setting was a floodplain, dissected from time to time by channels. Other fossils recovered from the locality include placoderms and a variety of other lobe-fins. Out of water, Tiktaalik could, at best, only have dragged its 2½-metre-long body. Monkfish, which like Tiktaalik have a wide mouth, flattened skull and eyes on top of the skull rather than on the sides, again alert us to other possibilities, for in addition to walking they use their fins to scoop out sediment from under their bodies and make a hollow, in which, having camouflaged themselves to match their surroundings, they lie in wait. At the right moment, they strike the prey by suddenly pushing with their fins upwards and forwards. The feeding strategy of Tiktaalik may have been similar. Several lineages of fish, such as mudskippers, rockskippers and blennies, have even evolved an ability to crawl out of water. However, there is no reason to think they are on the way to leaving the water altogether.
- The characteristics shared by the Devonian tetrapods and certain lobe-fins were those that equipped them for their shallow-water environment. They differed from each other because they were each optimised for their own particular life-style, not because they were evolutionarily on their way to somewhere else.
- The first known tetrapods with limbs and digits lived wholly (Acanthostega) or primarily (Ichthyostega) in the water, so the acquisition of such appendages does not support the story of how ‘our fishy ancestors began hauling themselves onto dry land’. Their splayed limbs were designed for paddling, not walking.
- There remains a large morphological gap between Tiktaalik and the first tetrapods.
- Trackways attesting the existence of tetrapods predate Tiktaalik by 10 million years, so Tiktaalik cannot be ancestral to the first tetrapods.
- How does one fill the gap between Tiktaalik, a fish with no legs, and an aïstopod such as Lethiscus, which within 20 million years of Tiktaalik had (it is thought) acquired legs and limb girdles, then lost them and ended up looking more like a snake than any tetrapod?
There is an odd sense of unease pervading the research community. It has been brought about by a head-on collision between an established interpretative scenario for the origin of tetrapods, seemingly well-grounded in the body fossil record, and two sets of tetrapod trace fossils from Poland and Ireland that appear to flatly contradict it.
originated no later than the beginning of the Middle Devonian, and coexisted as ecologically separated radiations for at least 15 million years; … that all Devonian and many later tetrapods … retained a permanent dependence on the aquatic environment, allowing only relatively brief terrestrial excursions; that this continued dependence explains the repeated evolution of ‘secondarily aquatic’ forms among them; and that all fully aquatic tetrapods, including Acanthostega, are likely to represent reversals from a somewhat more terrestrial ancestry.
In other words, the fossils of the Late Devonian were evidence of an earlier movement towards terrestrialisation that the fossil record fails to capture. Not only was that process never completed, but by the Late Devonian/Early Carboniferous it had gone into reverse. Consistent with the evidence of lepospondyls such as Lethiscus, Ichthyostega and Acanthostega represented a loss of terrestriality. One elpistostegalian fish of the Late Devonian itself shows evidence of ‘a secondary return to open water’ (Stewart et al. 2022).
In the light of Tiktaalik in particular, it seems likely that a few lobe-fins (amongst the huge range of species that arose in the Devonian) did evolve into aquatic tetrapods. Vegetated, fresh or brackish-water shallows were a new environment, and the acquisition of ‘tetrapod-like’ features was what fish needed to exploit them. The genetic programs that produced the innovations were engineered with the future environments in mind. But that marked the limit of the program’s ability to innovate in that direction. The alleged transition from aquatic to terrestrial tetrapods (i.e. amniotes) remains undocumented. Acanthostega and Icthyostega were both dead-ends.
One is tempted to say that the transition from fish to aquatic tetrapod happened only once, and that this itself is evidence that the evolution was pre-programmed. We do not, after all, see lobefins morphing into tetrapods at any later time in geological history. True, some rayfins – e.g. mudskippers, climbing perch – later evolved an ability to crawl on land and survive there for short spells, and other rayfins, following as it were in Acanthostega’s footsteps, evolved ‘tetrapod-like’ features that enabled them to walk under water. Some even transformed their fins into wings and learned to fly for short distances! But they too are dead-ends. Evolutionary biologists do not argue that mudskippers are on the way to becoming reptiles or that flying fish are on the way to becoming birds. Nor are newts and frogs about to abandon their watery origins. Toads have gone further than most, but even they must migrate to ponds to mate and spawn. They still start their lives as tadpoles.
Not that such a transition would have been impossible. Evolution does not happen by magic, and is not a process of random mutations fumbling in the dark. Genomes could have been programmed to go all the way, from fully marine to fully terrestrial. In the absence of any evidence for the transition, however, one must deduce that amphibians and amniotes had separate ancestries – what one might term the Noah’s Ark scenario. Aquatic tetrapods could not have been on the ark, since the ark preserved only air-breathing animals. Aquatic tetrapods therefore must have originated from fish in the sea, and could not always have been tetrapods since, before the Devonian, the environments to which they were adapted – swamps, lagoons, estuaries and the like – did not exist. Moreover, any antediluvian tetrapods would have been destroyed in the Cataclysm.
Unfortunately, the National Academy of Sciences is intent on blurring the differences. Tiktaalik, it says, had both the ‘features of fish (scales and fins) and features of land-dwellers (simple lungs, flexible neck, and fins modified to support its weight). The bones in the limbs of this fossil, named Tiktaalik, resemble the bones in the limbs of land-dwelling animals today.’ This is not a truthful statement. Lungs, simple or otherwise, were not mentioned in the scientific report. A mobile neck (‘flexible’ is too strong, for even Acanthostega did not have a flexible neck) might have helped the animal to gulp air, or even raise its head above water and target prey by the water’s edge, but it was not a uniquely tetrapod feature. The 2-metre-long lobe-fin Mandageria also had a mobile neck (an example of ‘parallel evolution’), as does the modern eel catfish. And Tiktaalik’s fins had no ability to support its weight out of water. Is it only by making misleading and partly false statements that the Darwinian story can be maintained?