







 – photo: Paul Strother") New land still forms today, and the way it forms is essentially the same as the way it formed in the Proterozoic and Palaeozoic. New seafloor was volcanically generated at mid-ocean ridges and, to compensate, old seafloor was subducted beneath continental plates. Water rising from the oceanic plate as it sank into the mantle lowered the melting point of the rock above it and caused magma to rise to the surface, erupting to form volcanic mountains such as (to cite relatively recent examples) the Rockies and Andes. These added mass to the continental margin, some of the mass eroded into the sea and combined with sediments that were being scraped off the subducting oceanic plate, and over time the area of the continental margin grew.
New land still forms today, and the way it forms is essentially the same as the way it formed in the Proterozoic and Palaeozoic. New seafloor was volcanically generated at mid-ocean ridges and, to compensate, old seafloor was subducted beneath continental plates. Water rising from the oceanic plate as it sank into the mantle lowered the melting point of the rock above it and caused magma to rise to the surface, erupting to form volcanic mountains such as (to cite relatively recent examples) the Rockies and Andes. These added mass to the continental margin, some of the mass eroded into the sea and combined with sediments that were being scraped off the subducting oceanic plate, and over time the area of the continental margin grew.
It took time for plant and animal communities to establish themselves on this barren land. Terrestrial animals, be they insects or more fleshy creatures, herbivores or carnivores, depend directly or indirectly on plants for their food, so we look at the plant record first and some of the evidence for the first stages in the process. Which better accounts for the fossil sequence: recolonisation or step-by-step evolution?
Initially, the process may not have been dissimilar to the colonisation of volcanic islands that form today, such as Surtsey, which formed off Iceland in the 1960s. By 1971, 83 vascular plants had become established as a result of seed dispersal by wind and water (Fridriksson 1975). Iceland itself is a relatively new land mass. Since land formation has never totally stopped, there are opportunities to observe the sequence of events from the moment the lava solidifies. Some eruptions produce completely new land; others resurface existing continental areas, but these too offer insights. Though modern eruptions are puny compared to the cataclysm that caused the entire planet to be resurfaced, it is evident that vegetation greens up the land in stages, not all at once.
Typically the first colonisers are microbes, then lichens and mosses, then herbaceous plants, shrubs and trees. Depending on climate and other factors, the later stages may never be reached, and successions do not always follow such a pattern. The order is determined by rates of growth and generation times, and by the fact that the later-arriving organisms depend for food on the earlier ones. Each stage builds on the one before, leading to a mature ecosystem where nutrients are recycled and a great variety of organisms interact to each other’s benefit.

Bacteria, fungi and algae are the first to build communities. They release chemicals which break the rock into grains. Carbon dioxide dissolved in rainwater creates a weak acid which reacts with the rock minerals, washing some away in solution and turning the residue into clay. Soil bacteria fix nitrogen into compounds which are useful to plants. Roots and humic acids break up the ground further and enhance the weathering process. In time, burrowing ants and earthworms join the ferment. Over months and years, progressively greater depths of rock are converted into fertile soils. The succession of events is ecological, not evolutionary. Micro-organisms do not evolve into mosses, or mosses into ferns. Each group has an independent origin from beyond the area colonised.
The correspondence between modern ecological successions and the order of fossils is not perfect. In important respects the yet-to-be-vegetated world was different from the modern world. Rivers in the sense of meandering long-lived low-energy channels cutting into a vegetated landscape did not exist; most were high-energy, and their courses continually changing. Alluvial landscapes in the Cambrian consisted of broad, unconfined sheets of coarse sediment; mud comprised less than 15% of the sediment and included little organic matter (Davies & Gibling 2010). Earthquakes and gravity slides were continually eroding the mountains and distributing coarse sands upstream and finer sand downstream. Flash floods were generated not so much by rain-storms as by pulses of water surging through faults, as they suddenly slipped and brought up water from beneath the crust. Plants could not establish themselves so long as surfaces were continually being buried.
In fast-flowing waters large particles settle out more readily than small particles. As conditions became quieter through the early Palaeozoic, the proportion of mudrock to sandstone increased, while the overall size of the sand grains decreased. Pauses between one depositional event and another lengthened, eventually becoming long enough for simple plants to establish themselves: plants that grew quickly and did not require a large investment of biological energy. Thus, for some while, plant fossils tended to represent only pioneer species, at the beginning of potential successions. Species that took successions beyond the pioneer stage only stood a chance of impressing the record when a locality had been stable for longer than a few years and was only then overtaken by a depositional event rapid enough for the vegetation to be fossilised.
The places where vegetation would most easily have become established were those furthest away from the highlands, such as low-lying plains and deltas. Here the energy of rainstorms and discharges of subterranean water was mostly dissipated. Tectonic processes in the mountains, coupled with the weathering effects of acid rain (at a time when there was much more carbon dioxide in the atmosphere), had already done the hard work of converting hard rock into grains. The successful plants were those suited to wetland environments, comparatively simple in design. They did not require mature soils. Disseminating their spores in the water, they reproduced quickly and proliferated over wide areas.
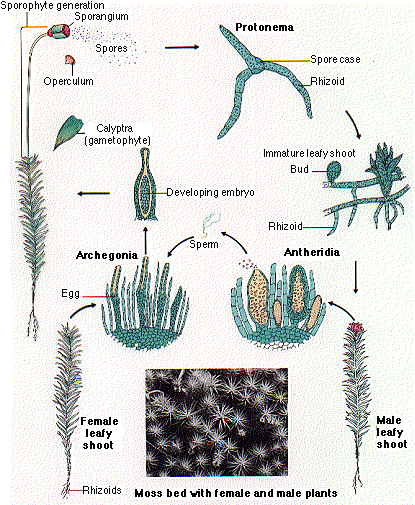
 Bacteria, fungi and algae reproduce by means of spores. So do certain plants, such as mosses, lycopsids (clubmosses), horsetails and ferns. Produced in organs called sporangia, plant spores have very resistant walls that protect them against desiccation, attack by microbes and damage during transport and, whether by wind and water, they are easily transported because of their lightness and smallness. Consequently they are not only the best preserved plant remains in old sedimentary rocks but also the first, and by far the most abundant. By contrast, the spores of bacteria, fungi and algae are rarely preserved.
Bacteria, fungi and algae reproduce by means of spores. So do certain plants, such as mosses, lycopsids (clubmosses), horsetails and ferns. Produced in organs called sporangia, plant spores have very resistant walls that protect them against desiccation, attack by microbes and damage during transport and, whether by wind and water, they are easily transported because of their lightness and smallness. Consequently they are not only the best preserved plant remains in old sedimentary rocks but also the first, and by far the most abundant. By contrast, the spores of bacteria, fungi and algae are rarely preserved.
Spores of mosses and liverworts are known from at least the mid Ordovician onwards, less certainly from the Cambrian, and their abundance increases with ascending stratigraphic level, as one would expect if plant fossils were reflecting the progressive recovery of vegetation. The earliest moss macrofossils are fragments from the Early Carboniferous and strikingly similar to extant peat mosses. Macrofossils before the Permian are scarce, presumably because conditions were unfavourable for their preservation, though just why remains unclear. The Ordovician-early Silurian in particular was a time of persistently high sea-levels, leaving few continental deposits. Many of the earliest spores occur in nearshore marine deposits, blown out to sea by the wind. It is of course easier to understand the delay in macrofossil preservation within a timescale that allocates only thousands of years to the greening process. According to conventional dating, the mid Silurian to the end of the Devonian occupies 70 million years.
Though simpler than most plant types, mosses and liverworts are intrinsically complex, as is apparent from their life cycle and their ability to photosynthesise. As with all plants and some algae, their generations alternate between haploid phases, which have one set of chromosomes, and diploid phases, which have two. The moss forms that we usually see are the haploid generation. The diploid phase is the short-lived result of self-fertilisation and grows on top of the plant, until it releases haploid spores and the cycle starts again. The free-swimming sperm require water for their fertilisation.
How this system came to be is unknown, and how mosses and liverworts fit into the evolutionary tree of life is unknown. Whether or not they are related to each other, they are neither preceded by evolutionary forerunners nor followed by evolutionary successors. Capable of withstanding huge fluctuations in moisture and temperature, they can survive in habitats inimical to other plants. They have changed little over time because they are so well-designed for their particular ecological role. That said, although unchanging as a group, they have become extremely diverse. Some 20,000 species (12,000 mosses, 8000 liverworts) are known today. Modern liverworts are thought to be less complex than their ancient ancestors (Puttick et al. 2018).
Mosses today are among the most common and ubiquitous eukaryotic organisms on the planet, so it is not surprising that they were among the first to appear in the fossil record. They are vital for maintaining plant diversity, soil microbial communities and multiple ecosystem services and are especially important for supporting such services where vascular-plant cover is low (Eldridge et al 2023). Not surprisingly, mosses were among the first plants to grow on Surtsey. Within a few years they were growing around crevices and holes where steam kept the volcanic rock damp. Nitrogen-binding cyanobacteria were also found around the steam-holes. Just seven years after the first eruption in 1963, mosses and lichens were widespread on the bare lava.
 Fungi are not plants but multicellular organisms (but yeasts are unicellular) in a kingdom of their own. As with the other kingdoms their origin is a mystery. More than 100,000 species have been described, representing perhaps 5% of the total. Some are aquatic, most terrestrial. They play a vital role as decomposers, recycling organic remains back to the environment in forms digestible by other organisms. Nearly all plants depend on symbiotic fungi called mycorrhizae to help the roots absorb minerals and water from the soil, while the fungi benefit by receiving carbohydrates from the plants.
Fungi are not plants but multicellular organisms (but yeasts are unicellular) in a kingdom of their own. As with the other kingdoms their origin is a mystery. More than 100,000 species have been described, representing perhaps 5% of the total. Some are aquatic, most terrestrial. They play a vital role as decomposers, recycling organic remains back to the environment in forms digestible by other organisms. Nearly all plants depend on symbiotic fungi called mycorrhizae to help the roots absorb minerals and water from the soil, while the fungi benefit by receiving carbohydrates from the plants.
Until recently the oldest traces of terrestrial fungi were filaments and spores dated to the Ordovician, about the same time as plant spores appear. They bear a strong resemblance to certain modern mycorrhizae that form filaments in or among roots as a way of increasing the surface area for absorption. Fungus-looking filaments also occur in the Silurian, some again looking like extant species. Numerous types of fungi have been observed among the plants in Scotland’s Rhynie Chert, dating to the early Devonian. In the Congo, shales interpreted as collecting in ephemeral ponds preserve the remains of fungi 300 million years older than even the Ordovician examples. Such records are prone to be broken, because the dates assigned reflect accidents of discovery and preservation, not the time at which a group can be said to have evolved. Being so delicate, fungi are rarely fossilised.
Galls caused by fungi appear almost as early as the affected plants themselves (Knecht et al. 2024), including, in the Carboniferous, galls on Calamites and Sigillaria stems, horsetails and marattioid ferns. (See ‘Rising Fast: The First Trees‘.)
Lichens consist of fungi living in partnership with cyanobacteria or algae – an even closer mutuality than between some fungi and roots. The fungus contributes to the partnership by taking up water and salts, the alga by producing the chlorophyll needed for making carbohydrates. Together they play an important role in breaking up bare rock into soil.
DNA analysis of extant forms leads to the conclusion that lichens arose independently five times or more. Their fossil record is poor, but cyanobacteria and algae first appeared in the Archaean, fungi probably in the Proterozoic, and there are reports of soils with lichen-like fossils dating back to the Cambrian. The oldest unequivocal lichen is Spongiophyton minutissimum, a widespread early Devonian fossil.
 Plant photosynthesis involves the absorption of carbon dioxide and the emission of oxygen, mediated through pores. Since large amounts of water vapour are lost in the process, plants higher than a few centimetres require specialised tissues to draw water from the ground. In vascular plants these tissues are of two types: xylem, which pipes up water and minerals from the roots to the leaves, and phloem, which distributes sugar and other products of photosynthesis from the leaves to the roots. Xylem also provides stems with rigidity, another requirement if plants are to grow higher than a few centimetres. The appearance of these tissues was of great significance, for they characterise most plants in the world, including low-growing ones such as some of the grasses. Some mosses also have conducting cells, but these originated independently from those in vascular plants. Like mosses and liverworts, vascular plants alternate between haploid and diploid phases, but in contrast to them, their dominant phase is the diploid, sporophyte phase.
Plant photosynthesis involves the absorption of carbon dioxide and the emission of oxygen, mediated through pores. Since large amounts of water vapour are lost in the process, plants higher than a few centimetres require specialised tissues to draw water from the ground. In vascular plants these tissues are of two types: xylem, which pipes up water and minerals from the roots to the leaves, and phloem, which distributes sugar and other products of photosynthesis from the leaves to the roots. Xylem also provides stems with rigidity, another requirement if plants are to grow higher than a few centimetres. The appearance of these tissues was of great significance, for they characterise most plants in the world, including low-growing ones such as some of the grasses. Some mosses also have conducting cells, but these originated independently from those in vascular plants. Like mosses and liverworts, vascular plants alternate between haploid and diploid phases, but in contrast to them, their dominant phase is the diploid, sporophyte phase.
How natural selection acting on genetic mutations leads to new designs is far from clear, and assuming it did, one would expect it to give rise to no more than adequate design. The characteristics of distinctly good design, such as energy efficiency or ingenuity in the solution of a given problem, are evidence of an intelligent creative power. In this instance the design is superb. ‘It is difficult to imagine a cheaper process for driving the transpiration stream,’ writes John Sperry. The coupling between evaporation at the plant surface and the negative pressure achieved by capillary forces in the cell walls produces the driving force almost free of charge. Xylem conductivity per area exceeds that of non-vascular plants by six orders of magnitude.
 The minimum requirement for such a system is a genetic program that causes the death of the cells lining the conduits and the manufacture and deployment of a substance (lignin) to strengthen the cell walls against collapse. These innovations were already in place by the late Ordovician, when vascular plants left evidence of themselves in the form of characteristic spores. The oldest known whole vascular plant, Cooksonia, did not appear until the mid Silurian, when the processes of erosion and deposition around them were presumably now less destructive. Cooksonia was small (less than 10 cm high), simple in appearance, and like the spores initially rare. By the end of the period (supposedly millions of years later) it was globally widespread, though still rare, and comprised at least five species.
The minimum requirement for such a system is a genetic program that causes the death of the cells lining the conduits and the manufacture and deployment of a substance (lignin) to strengthen the cell walls against collapse. These innovations were already in place by the late Ordovician, when vascular plants left evidence of themselves in the form of characteristic spores. The oldest known whole vascular plant, Cooksonia, did not appear until the mid Silurian, when the processes of erosion and deposition around them were presumably now less destructive. Cooksonia was small (less than 10 cm high), simple in appearance, and like the spores initially rare. By the end of the period (supposedly millions of years later) it was globally widespread, though still rare, and comprised at least five species.
The relationship of Cooksonia to the later vascular plants is as little understood as the origin of the whole group. This later vegetation included zosterophytes (from the Greek for garland, describing the successively arranged sporangia, and phyton, meaning plant), rhyniophytes (named after the Scottish village of Rhynie), trimerophytes (referring to their multiple branching, as illustrated by Psilophyton below) and lycopods (referring to the resemblance of some branch tips to a wolf’s paw). Most did not have leaves and roots, but the lycopod Baragwanathia had leaves (as do mosses), and all of them branched upwards off horizontally growing stems called rhizomes, which anchored the plants. In other respects they differed markedly from each other, notably in the design of their conducting walls, which were more complex and various even than those of extant plants. Since these diverse groups all appeared about the same time (late Silurian-early Devonian), a shared ancestry seems unlikely. The zosterophytes, rhyniophytes and trimerophytes died out in the course of the Devonian and left no descendants. A few lycopod species exist today, though much diminished both in size and diversity.

{kind=link}
Baragwanathia, the earliest examples of which come from Australia, is especially problematic because it is taller and more ‘advanced’ than its fossilised contemporaries. Other anomalously early appearances include vascular plant fossils from Late Silurian deposits on Bathurst Island, Canada (Kotyk et al. 2002), and Early Devonian deposits around Gaspé Bay, Canada. In the latter case, the plants achieved a stature of 2–3 metres and were fossilised in situ, showing that they were capable of rooting to a depth of nearly 1 metre (Elick et al. 1998). Despite similarities in anatomy (the presence of a root cap, root hairs and an endodermis), there is no evidence that the roots of lycopods and non-lycopods evolved from a common ancestor (Hetherington & Dolan 2018).
Because of these and many other discoveries the ‘somewhat simplistic’ picture of plant evolution has had to change (Edwards & Richardson 2004). Simplicity of outward form masks complexity at the cellular level. Almost from the moment they appear, vascular plants are diverse, making it difficult to see how, invisible to the fossil record, they might all have arisen from a single vascular plant ancestor. The gap between vascular and non-vascular plants such as the mosses and liverworts is even larger. So is the gap between all land plants and their closest presumed relatives, the green algae (Kenrick 2000). By contrast, the time gap between the vascular and non-vascular plants is comparatively small. Mega-evolution simply is not an appropriate inference to draw.
While it is recognized that the assemblages provide the most complete and extensive record of the history of vascular plants in a restricted geographical area during the time interval, it seems likely that major evolutionary innovation occurred elsewhere.
‘Major evolutionary innovation’ always occurs off camera. What we actually see is organisms that have already been ‘innovated’ – one might say, created – parachuting in from somewhere else, colonising virgin territory. Currently, vascular plants appear to have originated in Gondwana, i.e. present-day India and the southern continents (Anderson et al. 1999, Wellman et al. 2013).
That, perhaps, is where Noah’s ark was, a small enclave of human cultivation in the midst of what was now a huge supercontinent. New stocks of vegetation for replanting the Earth were not, for the most part, ‘just round the corner’. In the case of Surtsey, plant seeds were brought over in the gizzards of migrating birds, but birds in the Palaeozoic were scarce – none from the Palaeozoic have been preserved as fossils, and birds anyway are rarely fossilised – and many of the plant species characteristic of modern ecosystems had not yet evolved. On an earth initially denuded of fauna and flora there was little possibility of bringing in supplies from anywhere near. Some spores and pollen may have survived the Cataclysm and germinated in places unknown to us, but in general the planet had to be restocked from the area around the ark, where Noah’s descendants cultivated the ground using the fodder stored on board and manure produced by the animals. If the original garden of Eden had been a solitary enclave amidst an earth that was otherwise empty of vegetation – ‘for no bush of the field was yet on the earth and no plant of the field had yet sprung up’ – so now was this little oasis, this ancient equivalent of Kew Gardens’ Millennium Seed Bank.