








The rise of plants in the Devonian was a step change. In addition to the mosses and marsh vegetation of the Silurian, now every major plant type except flowering plants was present. Extraordinarily complex organs and tissue systems appeared as if from nowhere. It was a ‘novelty radiation’ rather than a slow accumulation of infinitesimally small changes: stems with an intricate fluid transport mechanism, lignin, structural tissues, pores for controlled gas exchange (stomata, although some mosses have stomata on their sporangia), leaves and roots of various kinds, seeds.
Almost from the first, the plants were not isolated pioneers but members of small, mixed communities, fledgling ecosystems in which springtails, trigonotarbids, insects, centipedes and millipedes played their part. Some of the animals were carnivores, but they were not competing in overcrowded environments where only the fittest survived, but living in balance with each other, interdependently. Species numbers were initially low.
The oldest known fossil forest is that of the Plattekill and Gilboa Formations, New York State, dating to the Middle Devonian. The Gilboa forest comprises two horizons each 70– 90 cm thick, separated by 2.5 m of fine deltaic sandstone. Each horizon represents little more than a single generation, and the 4 m in total probably no more ") than half a century. The fossils present a dramatic contrast to the ground-hugging plants of earlier times. Although the new forms still grew in a water-logged soil, they were full-grown trees, their tapered pith-filled trunks rising to a height of 9 metres or more. The tallest was the fern-like Eospermatopteris. Its bulbous base gave it a low centre of gravity, anchored by thin, shallow, unbranching roots that radiated outwards and downwards. There were also smaller trees and shrubs.
than half a century. The fossils present a dramatic contrast to the ground-hugging plants of earlier times. Although the new forms still grew in a water-logged soil, they were full-grown trees, their tapered pith-filled trunks rising to a height of 9 metres or more. The tallest was the fern-like Eospermatopteris. Its bulbous base gave it a low centre of gravity, anchored by thin, shallow, unbranching roots that radiated outwards and downwards. There were also smaller trees and shrubs.
{kind=link}
The tree habit appeared independently in several other lineages – lycopsids, progymnosperms, horsetails – and all but the horsetails achieved this in the interval in the Middle to Late Devonian. This puzzling phenomenon where Nature seems to invent the same feature more than once is termed ‘convergence’ or parallel evolution. It is pervasive in large-scale phylogenies, occurs amongst all types of organism, and characterises even the most complex innovations. Photosynthesis is another example: a highly engineered process that occurs in bacteria, algae and protists as well as plants – entirely separate biological kingdoms. Sporopollenin occurs in the cell walls of some green algae and is the main polymer forming the tough outer walls of plant spores. Within plants, major innovations such as stomata, sporangia, xylem, leaves, roots and wood tissues all arose more than once. Leaves arose at least five times, perhaps as many as nine. Wood tissue – involving either a unifacial or a bifacial cambium – arose at least seven times.
As Michael Donoghue noted, ‘recent phylogenetic findings are making it increasingly difficult to sustain the traditional view’ whereby the identification of key innovations is the way to distinguish new branches in the tree of life. If it is difficult to believe that Nature could engineer these fantastically complex innovations even once unless pre-programmed to do so, how much more incredible is it that the same innovation could arise a second time, or a third or a ninth. Convergence is evidence that the history of evolution cannot be reduced to a single phyogenetic tree. The structures in question were products of design, not accident.
Long, foreign-sounding names and complex biology can make the plant world of the time seem alien. Here we offer a brief guide through the jungle.

Progymnosperms, the earliest-known trees, first appeared in the Middle Devonian and became extinct in the Early Carboniferous. The innovation of a bifacial cambium enabled them to produce wood (secondary xylem) and thereby grow outwards as well as upwards, in the same way as trees do today. Unlike the later-appearing gymnosperms, to which they were probably ancestral, they reproduced by means of spores rather than seeds. The relationships between the three progymnosperm orders are unclear.
Archaeopteris, the best known example, was the first modern tree. Being only slightly younger than Eospermatopteris, it was virtually the first tree. It had an extensive root system, produced leaves, grew to a height of 30 metres or more, and had laterally growing branches. A swelling at the base of the branches formed a supporting collar, while layers of wood dovetailed into the trunk to give additional strength. In one respect Archaeopteris was not modern, for today large woody trunks are produced only by seed plants. Its spore-system of reproduction meant that it needed to be close to bodies of ponded or flowing water, allowing sperm to swim to the egg. Quite possibly these were the only widely colonisable environments in existence at this time, in which case it would make sense that the first trees to appear were those adapted to such environments.
Diversification led to several species of Archaeopteris. Occurring in large numbers on every continent, they colonised floodplains and coastal areas on a global scale. Along with the much less widespread Eospermatopteris they created the first forests. Forests ceased on this scale in the wake of events causing the mass extinctions at the end of the Devonian and did not recur until well into the Carboniferous, when several new types of woody tree appeared.
 Lycopsid is the name for lycopods (clubmosses) and related forms. The informal name refers to their moss-like appearance, though they are not true mosses, and to the club-shaped sporophyll clusters at the tips of their tiny branches. (A sporophyll is a leaf which bears the capsule in which spores develop, called a sporangium.) Only a few lycopod genera exist today, and they look diminutive compared to their largest forebears.
Lycopsid is the name for lycopods (clubmosses) and related forms. The informal name refers to their moss-like appearance, though they are not true mosses, and to the club-shaped sporophyll clusters at the tips of their tiny branches. (A sporophyll is a leaf which bears the capsule in which spores develop, called a sporangium.) Only a few lycopod genera exist today, and they look diminutive compared to their largest forebears.
Lycopsids are distinguishable by their sporophylls and their spirally arranged microphylls (small spine-like leaves with just one vein). The oldest known lycopsid is Baragwanathia, which we mentioned in Colonising wetlands. It first appeared in the Late Silurian and survived into the Early Devonian. Its leaves were separate from the sporangia and not fully veined. Another Early Devonian lycopod, Drepanophycus, is almost indistinguishable from the still extant shining clubmoss, Huperzia lucidula, a rare case of stasis through the group’s entire record.
Within the lycopsids were the Protolepidodendrales, a widespread order whose leaf-tips were forked. Some grew to a height of 50 cm or more. They may have been ancestral to the arborescent (tree-forming) lycopods of the Late Devonian and Carboniferous.
Arborescent lycopods differed most notably from that group in having a bipolar shoot system (with branches growing upward and roots downward) and growing erect rather than along the ground. In these respects the Middle Devonian Longostachys has something of an intermediate character. Only 1.5 metres tall, it had a slender trunk that branched at the top to form a crown of narrow branches and terminal cones. At the base it had downward-branching roots. Arborescent lycopods also had rootlets that grew perpendicularly and radially off the rooting organs, like the hairs of a bottlebrush. How they evolved remains a mystery: their appearance in the record is abrupt (Gensel & Berry 2001).
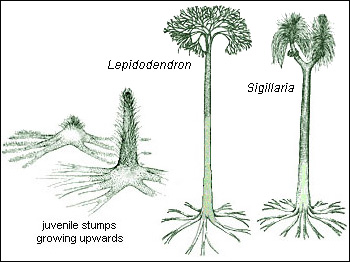
Arborescent lycopods such as Sigillaria and Lepidodendron were the major components of the vast Carboniferous swamp forests that ended up as coal seams. These trees produced almost no woody tissue. Instead the trunk was filled with pith and strengthened by a thick cortex, which extended from the cambium outwards to a bark-like, decay-resistant periderm. Long grassy leaves grew from the trunk, and following leaf-fall the scars on the leaf cushions composed a distinctive geometric pattern. In contrast to the brown, non-photosynthetic stems of modern trees, photosynthesising bark covered the whole trunk, a super-efficient energy factory that enabled the trees to reach full height (30 metres or more) in just a few years. Only towards the end of their lives did they form lateral branches and put out a leafy crown, and that was in order to reproduce before they died, by means of spore-bearing cones. Because of their pillar shape, they were able to grow in great density, up to 800 trees per acre.
") Their rooting systems, called stigmaria, were designed to float. They supported the lightly-built trees by splaying out horizontally, and the collapsed state of the fossils (or sediment infill where not collapsed) shows that they were hollow, or at best pithy, and immersed in water. Spiky appendages around the stigmaria radiated bottlebrush-like in every direction and were also hollow. From time to time they were shed and left scars, just like the leaves. Not having an anchoring function, they probably photosynthesised like the leaves. The stigmaria were incapable of penetrating clay soils and were not therefore rooted in ‘seat earths’, as is usually assumed. The clays, sandstones and even limestones of the sediments underlying the coals accumulated before the floating forests sank.
Their rooting systems, called stigmaria, were designed to float. They supported the lightly-built trees by splaying out horizontally, and the collapsed state of the fossils (or sediment infill where not collapsed) shows that they were hollow, or at best pithy, and immersed in water. Spiky appendages around the stigmaria radiated bottlebrush-like in every direction and were also hollow. From time to time they were shed and left scars, just like the leaves. Not having an anchoring function, they probably photosynthesised like the leaves. The stigmaria were incapable of penetrating clay soils and were not therefore rooted in ‘seat earths’, as is usually assumed. The clays, sandstones and even limestones of the sediments underlying the coals accumulated before the floating forests sank.
Most of the arborescent lycopods went extinct before the end of the Carboniferous as their watery habitats dried out. Growing alongside them was a related group of smaller mostly unbranched forms, called the isoetaleans. These appeared about the same time as the arborescent forms, in the Late Devonian, but were also affected by extinctions. Today the order Isoetales is represented by only a single genus, the tiny semi-aquatic Isoetes (quillworts).
Six major groups are distinguished under this very broad term: whisk ferns, ophioglossoid ferns, cladoxylopsids, marattioid ferns, leptosporangiate ferns and horsetails. Although the first two groups may be related, relationships among the others remain elusive (Pryer et al. 2004). Ferns in the broad sense probably do not constitute a single-ancestor group (Rothwell & Nixon 2006). Cladoxylopsids, horsetails, leptosporangiate ferns and marattioid ferns are informally known as tree ferns. After most arborescent lycopods went extinct, these – but not cladoxylopsids, which went extinct in the mid Carboniferous – became the dominant plant group. Some North American coals from the Late Carboniferous consist up to 75% of tree fern remains.
Whisk ferns have no fossil record. Because of their simple body plans (for example, no roots, and highly reduced leaves), they were long thought to be related to some of the earliest vascular plants, such as Rhynia. However, most experts now believe they derive from more complex ferns that did have roots. They are an example of evolution proceeding in the reverse direction, being ‘advanced’ in time but comparatively simple in form. The similarity with the earliest vascular plants, while thought-provoking, is fortuitous. Ophioglossoid ferns go back no further than the earliest Cenozoic.
The earliest fern-like plants to appear were the cladoxylopsids, in the Middle Devonian, some of which grew to tree height. Although only recently identified as such, the most famous of the cladoxylopsids is the tree whose stumps thronged the Gilboa forest, Eospermatopteris. It photosynthesised through fronds that spread out from the crown like a feather-duster. As the trunk grew upwards, the branches dropped off and the outer tissues of the trunk remodelled themselves into longitudinal strands, so that the branch scars gradually disappeared. Present-day cycads and palms have a similar structure, but are seed plants (see below).
 Horsetails are characterised by grooved, jointed, hollow stems, whorls of highly ‘reduced’ leaves at the joints, extensive underground rhizomes, and sporangium-bearing stalks that cluster into cone-like ‘strobili’. Horsetails go back to the Late Devonian and rapidly diversified. They already comprised three orders when they peaked in abundance in the Late Carboniferous. Like the arborescent lycopods, they grew as tall as trees, and produced secondary tissue only on the inside of the cambium. Today they are represented by just one genus, Equisetum, and are diminutive by comparison with their forebears.
Horsetails are characterised by grooved, jointed, hollow stems, whorls of highly ‘reduced’ leaves at the joints, extensive underground rhizomes, and sporangium-bearing stalks that cluster into cone-like ‘strobili’. Horsetails go back to the Late Devonian and rapidly diversified. They already comprised three orders when they peaked in abundance in the Late Carboniferous. Like the arborescent lycopods, they grew as tall as trees, and produced secondary tissue only on the inside of the cambium. Today they are represented by just one genus, Equisetum, and are diminutive by comparison with their forebears.
Leptosporangiates are the most diverse of the fern groups. Of the dozens of families classified, one – the polypods – accounts for about 60% of the 11,000 present-day species, due to an extraordinary burst of diversification from the Cretaceous onwards. (This would be less anomalous in relation to the proposed, re-calibrated timescale which lengthens towards the present.) Leptosporangiate ferns appeared in the Early Carboniferous and soon after diversified into six or more lineages, none of which are easily traceable to present-day ferns.
Marattioid ferns first appeared in the Middle Carboniferous with later species reaching heights of up to 8 metres. Unlike most arborescent plants, they did not produce secondary xylem. Instead, the trunk was encased by a dense mantle of intertwining roots. Extant representatives of this group look much the same as their fossil ancestors.
As with the other ‘ferns’, there are earlier plants (such as Sphenophyllum, Hyenia) that although hinting at a relationship with the horsetails have so far resisted attempts to classify them within an orderly evolutionary scheme. The Late Devonian Pseudobornia reached heights of up to 20 metres.
, from Craigleith, now outside the Natural History Museum")
Seed plants include both the gymnosperms – plants with naked seeds – and the angiosperms, the ‘flowering plants’, whose seeds are enclosed within a fruit. Angiosperm fossils date back no further than the early Cretaceous, long after the gymnosperms first appeared, and probably descended from a gymnosperm lineage. Angiosperms may have descended from one or more gymnosperm lineages, but if so, the path from one to the other remains unclear. In this respect, the explosive innovation of coloured petals, stamens, carpels, scent and nectar in the Cretaceous remains the ‘abominable mystery’ that it was for Darwin. The appearance of already diverse non-flowering seed plants in the Devonian is just as difficult to understand (Pretianni & Gerrienne 2010).
Seeds are analogous to the amniote egg. They consist of an embryo and a store of food (analogous to the yolk), surrounded by a protective coat (analogous to the shell). The advantage of this mode of propagation was that the male cells – the pollen – were transported to the ovule by wind rather than water, and fertilisation took place within the ovule. In addition, less surface water was required after seed dispersal for the plant to germinate. The food store enabled the embryo to establish itself in its new environment unaided, before having to draw nutrients and water from the soil.
Seed plants could thus colonise a wider range of environments than the vascular sporing plants. While gymnosperms, like other types of plant, may have mostly preferred the wetter parts of the Palaeozoic landscape, they also spread into drier though still fairly low-altitude locations. The photograph shows one of the giant trees recovered from Lower Carboniferous sandstones near Edinburgh and displayed outside the Natural History Museum in London. These trees had been plucked from their inland environment, transported by strong currents and buried where the sediments, as they settled, fanned into the sea.
Gymnosperms are no longer thought to correspond to a group traceable to a common ancestor (the first seed plant) from which all such organisms arose. The evolutionary pattern is complicated.
The oldest known seed plant is Runcaria, from the Middle Devonian. It displayed ‘a highly derived morphology compared to that of sporangia in progymnosperms’ and was ‘already complex’ (Gerrienne et al. 2004), indicating that seed plants must have originated earlier than the time the progymnosperms first appeared. This is also the implication of the Late Devonian seed plants at Elkins, West Virginia, and Taffs Well, near Cardiff, which reveal greater complexity and variation than expected at this stage in seed-plant evolution.
 Yet another group that has not conformed to expectations is the ‘seed ferns’. These used to be considered transitional between ferns and seed plants, but are now recognised to be a very heterogeneous group, with relationships among themselves no clearer than their overall relationship to non-seed-fern plants. The earliest seed ferns were mostly liana-like plants and vines from the Late Devonian. Examples from the Carboniferous and Permian include erect trees. Seed ferns finally became extinct in the Eocene.
Yet another group that has not conformed to expectations is the ‘seed ferns’. These used to be considered transitional between ferns and seed plants, but are now recognised to be a very heterogeneous group, with relationships among themselves no clearer than their overall relationship to non-seed-fern plants. The earliest seed ferns were mostly liana-like plants and vines from the Late Devonian. Examples from the Carboniferous and Permian include erect trees. Seed ferns finally became extinct in the Eocene.
Other distinct seed-plant groups without obvious ancestry include the cordaites, cycads and conifers. These emerged in the Late Carboniferous. Some cordaites were shrubs; others reached tree height, with narrow parallel-veined leaves up to 1 metre long. They died out at the end of the Palaeozoic. The cycads and conifers are of course more familiar, having descendants in the modern world.
Why is it, then, that we talk about the ‘Cambrian Explosion’ but not about a ‘Devonian Explosion’? The main reason is that the Cambrian was when nearly all animal phyla first appeared and there are many more animal than plant phyla. Most animal phyla consist of marine organisms, a few of which subsequently colonised the land and/or fresh water (e.g. snails). Heterogeneous phyla such as arthropods and chordates (vertebrates), both of which first appeared in the Cambrian, really consist of several non-related groups, and some of them are exclusively terrestrial. These did not appear until the Devonian, but because they are lumped with the phyla that appeared in the Cambrian, their sudden appearance does not seem quite so revolutionary.
Some basic body plans are defined by gaps at a lower level of classification than phylum: for example, the gaps between mosses and trees, or between fishes and the earliest tetrapod. Plants and animals appeared on land in the Devonian just as suddenly as they did in nearshore marine environments in the Cambrian. It was a ‘novelty radiation’, where new biological types appeared, not by evolution from organisms visible earlier in the fossil record, but from off-stage, where they were previously too few and too sparse to be fossilised.
Despite the repeated flooding and burial of surfaces that might have become soils, the order in which plants appeared in the fossil record was the order of ecological succession:
| Organism type | First appearance |
| Mosses | Cambro-Ordovician |
| Fungi | Ordovician |
| Small herbaceous plants | Late Silurian |
| Shrubs, bushes | Early/Middle Devonian |
| Trees | Middle Devonian |
Little by little, plants were being given more time to establish themselves in the same spot and develop vertically tiered communities. It was not mega-evolution Darwin-style.
The increasing diversity of plants reflected the increasing availability of diverse habitats. Owing to the declining rate of radioactivity (the main driver of plate tectonics), geological processes such as sea-level change, volcanism and mountain-building were slowing down and environments were gradually becoming more stable, allowing a variety of pre-existing plant types to begin exploiting them. At the start of the Devonian terrestrial environments were still very wet – not so much because of heavy rainfall as because a lot of water was still streaming up through the crust, from what remained of the ‘great deep’. By the end of the period braided and meandering channels – one can hardly call them rivers – were alternating with floodplains. Over time, as the Carboniferous unfolded, many of these landscapes further stabilised into organic-rich systems with alluvial islands between the streams (Davies & Gibling 2010, 2011).
Diversification within the plant types partitioned the landscape into ever smaller ecological niches. Channel-dissected mudflats began to fill with lycopsids and fern-like plants. Floodplains further inland became sufficiently stable for trees to reach maturity. Opportunist species of seed fern spread from these wetlands to exploit areas of greater stress and variability. Plants producing a large number of small seeds preceded those producing a smaller number of large seeds.
 The diversifying plants created new habitats as well as simply colonising them. Increasingly penetrative root systems helped to break up rocks and develop soils, which in turn promoted the growth of a greater range of plants. Vegetation helped stabilise land surfaces and river banks by reducing erosion. Forest canopies moderated humidity and temperature, allowing smaller plants to grow beneath them and providing shade for a variety of invertebrates. They also produced copious amounts of spores and leaf litter. Whatever was not eaten by invertebrates was decomposed by microbes and fungi. Soils enriched by herbivores and decomposers enabled the growth of other plant types, and other invertebrates preyed on the herbivores. In time a complex detritus-based food web began to develop, supporting in the drier parts of forests insects, mites, spiders, myriapods and the like, in the wetter parts tetrapods, molluscs, eurypterids and shallow-water fish.
The diversifying plants created new habitats as well as simply colonising them. Increasingly penetrative root systems helped to break up rocks and develop soils, which in turn promoted the growth of a greater range of plants. Vegetation helped stabilise land surfaces and river banks by reducing erosion. Forest canopies moderated humidity and temperature, allowing smaller plants to grow beneath them and providing shade for a variety of invertebrates. They also produced copious amounts of spores and leaf litter. Whatever was not eaten by invertebrates was decomposed by microbes and fungi. Soils enriched by herbivores and decomposers enabled the growth of other plant types, and other invertebrates preyed on the herbivores. In time a complex detritus-based food web began to develop, supporting in the drier parts of forests insects, mites, spiders, myriapods and the like, in the wetter parts tetrapods, molluscs, eurypterids and shallow-water fish.
The environments in which the coal-producing forests grew were shallow coastal basins that extended far inland. The submerged accumulations of litter so produced could reach great thicknesses. Within them palaeobotanists have been able to trace repeating successions of coalified flora, each recording a drying-upwards cycle as the water beneath the forest shallowed and the mat-forming lycopods were joined by more terrestrial species. The water-level rose and fell as the fault-bounded basins rose and fell. In time storms or tsunamis drowned the forests, bringing the cycle to an end. Later in the Carboniferous the oscillations weakened and the forests became drier, attracting a different mix of plant species and a variety of animals.